Molecular Cell | 何跃辉团队揭示二价染色质修饰及其阅读蛋白驱动染色质状态转变的分子机制
染色质修饰与重塑对于真核生物基因表达起着重要的调控作用。“春化作用”是植物中经典的染色质修饰介导环境信号响应的表观遗传调控范例。在十字花科模式植物越冬拟南芥的春化过程中,冬季的持续低温导致成花转变的抑制基因FLC的表达被沉默,春季升温后,这种沉默状态被稳定维持,赋予“冬季低温的表观遗传记忆 (epigenetic ‘memory of winter cold’)”,从而使越冬植物获得在春天开花的潜能。冷处理之前,FLC位点的染色质处于高度激活状态,富含histone 3 lysine-4 trimethylation(H3K4me3)、H3K36me3等转录激活性组蛋白修饰,冬季长期低温的处理诱导该位点的染色质转变为沉默状态,即形成多梳沉默域(Polycomb domain),富含转录抑制性的。然而,激活状态的染色质转变为沉默状态的调控机理仍不清楚。
2023年3月14日,北京大学现代农业研究院、生命科学联合中心何跃辉研究组在Molecular Cell在线发表了题为A pair of readers of bivalent chromatin mediate formation of Polycomb-based ‘memory of cold’ in plants的研究论文,报道了拟南芥中两个植物特有的二价组蛋白标记阅读蛋白识别FLC位点特定调控区域的二价染色质,在春化过程中驱动多梳沉默域形成的分子机制。

长时间持续低温(寒冬)诱导Polycomb Group (PcG)染色质修饰因子催化FLC位点的H3K27me3修饰。此前的研究发现,FLC基因第一个外显子和内含子的连接区域附近约500bp的范围为“Polycomb沉默成核区 (Polycomb nucleation region)”【1】。何跃辉团队早期的研究发现成核区内47-bp的 “低温记忆元件(cold memory element,CME)” 介导了FLC位点“冬季低温记忆”的形成【2】。本项研究发现CME所处的染色质区域,在春化过程中一直处于H3K4me3和H3K27me3共存的二价修饰状态,该区域的二价染色质可以被两个植物特有的同源组蛋白标记阅读蛋白EBS(EARLY BOLTING IN SHORT DAYS) 和SHL(SHORT LIFE)识别。EBS和SHL由识别H3K4me3的PHD结构域和识别H3K27me3的BAH结构域组成【3,4】。之前的结构生物学研究发现,由于BAH结构域与PHD结构域距离太近,单个EBS或SHL蛋白无法同时结合H3K4me3和H3K27me3,因此其结合二价染色质的能力未定 【4,5】。本项研究发现EBS和SHL可以形成同源或异源二聚体,从而识别CME区域的二价修饰。此外、EBS和SHL还与识别CME序列的DNA结合蛋白VAL1互作,协同招募冷特异的H3K27甲基转移酶复合体VIN3-PRC2至CME区域,促进H3K27me3修饰在该区域的富集。升温后,EBS、SHL、PRC2等扩散至整个FLC位点,催化并持续维持H3K27me3修饰;CME区的二价染色质和基因体其它区域的H3K27me3共同构成FLC位点稳定的多梳沉默域(Polycomb domain),赋予“冬季低温的表观遗传记忆”(图1)
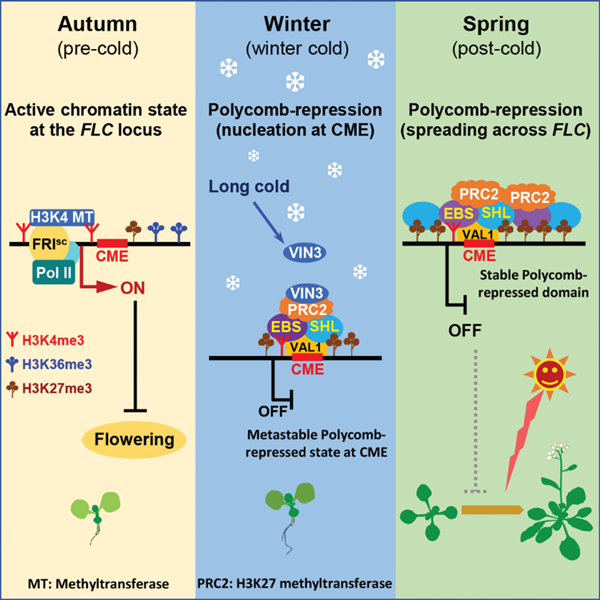
图1. 春化过程中EBS和SHL识别FLC位点CME区域的二价染色质,驱动多梳沉默域的形成,从而抑制FLC表达。
综上所述,该研究揭示了CME区域的二价修饰及其阅读蛋白EBS和SHL驱动了冷诱导的FLC染色质状态的转变,揭示了“冬季低温记忆”形成的分子与表观遗传调控机制,并为植物响应环境信号而转变二价染色质位点的修饰状态,并改变基因表达的调控模式提供了范例 (paradigm)。
北京大学、生命科学联合中心何跃辉教授为该论文的通讯作者,北京大学现代农学院博士后高政和中国科学院分子植物科学卓越创新中心的助理研究员李亚肖为共同第一作者。该研究得到了国家自然科学基金,生命科学联合中心、蛋白质与植物基因研究国家重点实验室以及中国科学院相关经费的资助。


