种子是植物生命的起点,能否在合适的时机萌发直接决定了植物能否抢占生长先机、躲避逆境。种子萌发受到休眠与萌发之间的精妙平衡调控,而活性氧(ROS)长期以来被认为是打破休眠、启动萌发的关键信号。然而,ROS信号究竟如何被传递到转录机器上,从而激活萌发相关基因的表达,这一核心问题一直悬而未决。

2026年5月27日,生命中心/清华大学生命学院方晓峰课题组在《高级科学》杂志(Advanced Science)上发表了题为《活性氧以来的分子伴侣通过调节GBF1凝聚体性质调控拟南芥种子萌发》(Redox-Dependent Chaperoning of GBF1 Condensates Regulates Seed Germination in Arabidopsis)的研究论文。该研究发现,转录因子GBF1通过液液相分离形成液态凝聚体,结合并抑制萌发促进基因CathB3的表达。而GIP1蛋白作为一类特殊的分子伴侣,能够调控GBF1凝聚体的大小和流动性,维持其高效的功能状态。当种子吸胀后,ROS爆发氧化GIP1,使其丧失伴侣活性,导致GBF1凝聚体流动性下降,解除对CathB3的抑制,从而启动萌发。这一“ROS-分子伴侣-转录凝聚体”轴线,首次将氧化还原信号与转录调控凝聚体的物态变化直接联系起来(图1)。
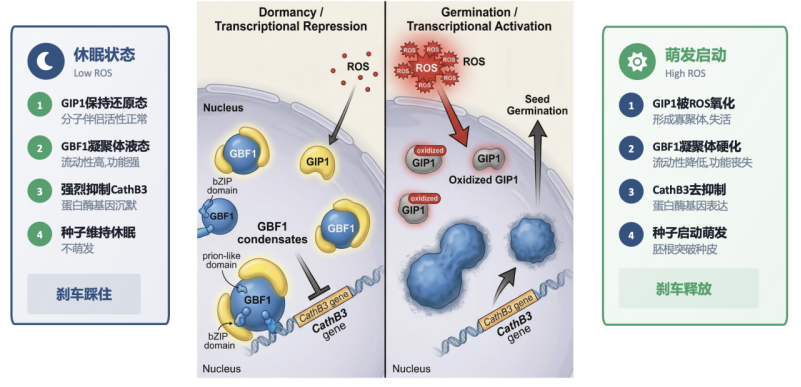
图1: GBF1-GIP1模块响应ROS调控种子萌发的工作模型(由AI生成)。
研究者发现体外纯化的bZIP转录因子GBF1蛋白在生理条件下形成规则的球形凝聚体,且具有快速荧光恢复能力和融合行为,呈现典型的液态特征。在体内,转基因拟南芥种子胚细胞核中,GBF1形成清晰的点状凝聚体。这首次证明GBF1在种子胚细胞核内发生相分离(图2)。随后研究者发现GBF1相分离可以促进DNA结合并保持转录活性。
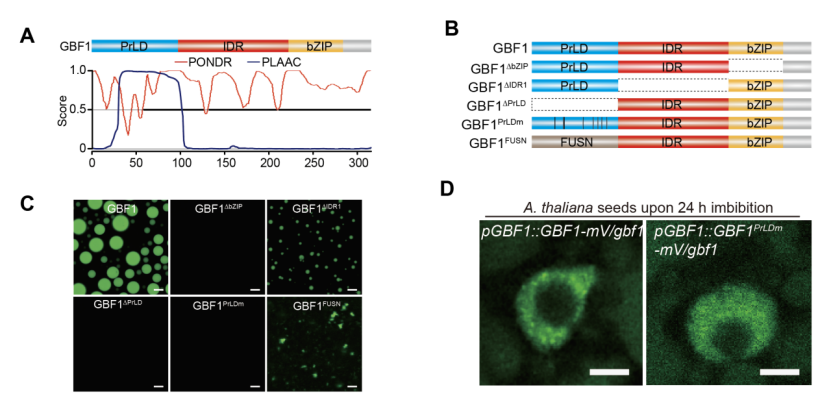
图2: bZIP 转录因子GBF1在体内和体外发生相分离。A) PONDR和PLACC算法预测GBF1相分离潜力。B) GBF1及其变体的蛋白结构域示意图。C) GBF1及其变体的体外相分离代表性图像。D) GBF1及其变体在拟南芥种子胚的聚集情况。
GIP1是GBF1的互作蛋白,在萌发种子胚中高表达,gip1突变体表现为萌发提前。遗传学分析表明,GIP1位于GBF1上游调控CathB3表达。有趣的是,GIP1本身不发生相分离,但能被招募到GBF1凝聚体中,并显著减小凝聚体的尺寸和数量。在体内,gip1突变体背景下GBF1凝聚体对流动性溶解处理更不敏感,表明其趋于固态(图3)。重要的是,液态GBF1凝聚体比老化后的固态凝聚体具有更高的DNA结合效率。因此,GIP1的伴侣功能在于维持GBF1凝聚体的液态和动态特性,防止其过度生长或固化,从而确保高效的转录抑制。之后研究者们发现ROS通过氧化GIP1的半胱氨酸、减少其伴侣活性,从而解除GBF1对CathB3的抑制,促进萌发。
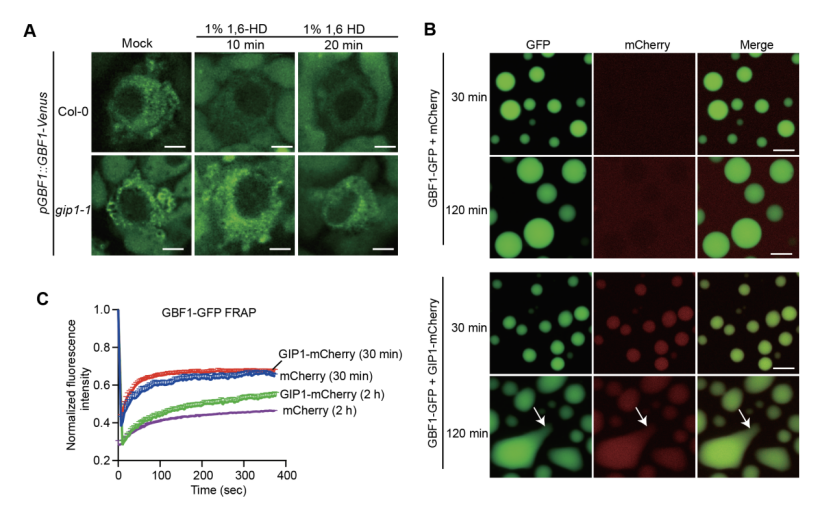
图3: GIP1调控GBF1凝聚体的大小和流动性。(A)野生型与gip1突变体种子胚中GBF1凝聚体对1,6-己二醇处理的敏感性。(B)有无GIP1时GBF1-GFP凝聚体的形态。(C)有无GIP1时GBF1-GFP凝聚体的流动性分析。
该研究首次揭示了一条“ROS信号→分子伴侣氧化失活→转录凝聚体固化→靶基因去抑制→萌发启动”的完整信号链。这一发现不仅解释了种子萌发的精确调控机制,也为作物抗逆育种提供了新的分子靶点,具有重要的理论意义和应用价值。
清华-北大生命科学联合中心研究员、清华大学生命学院副教授、绿色生物制造全国重点实验室研究员方晓峰为该文通讯作者。清华大学生命学院博士后王赟颖为第一作者。该研究得到了国家自然科学基金、科技部重点研发计划的资助。
原文链接:https://advanced.onlinelibrary.wiley.com/doi/full/10.1002/advs.202520599


